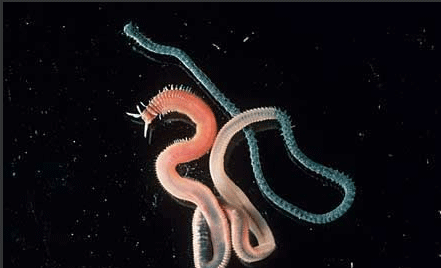
Annelid: The official worms
Worms do not comprise a single taxonomic category, but rather include members of quite different phyla such as Annelida, Platyhelminthes and Nematoda. Almost any invertebrate animal that is long and thin is called a "worm". We have even referred to ancestral organisms more resembling planula as "worms". Even caterpillars and beetle larvae are sometimes given that name.
Annelida is a group commonly referred to as segmented worms, descendants from ancestors existing perhaps well before the Cambrian. They are found worldwide from the deepest marine sediments to the soils in our city yards. There are about 13,000 species of annelids. They vary in size from only a few millimeters in length to the giant earthworms of Australia that may be three meters long.
Through most of the 20th century Annelida was split into three major groups; Polychaeta, Oligochaeta (earthworms etc.) and Hirudinea (leeches). Earthworms and leeches are the familiar annelids for most people, but polychaetes comprise the bulk of the diversity of Annelida and are found in nearly every marine habitat. It is now recognized that Oligochaeta and Hirudinea, comprised of several thousand species, form a clade and should be referred to the Clitellata. Polychaetes are less related to either living members of this group, although they are the oldest line of still existing Annelids.
Much discussion surrounds the origins and homology of segmentation, which is a characteristic shared not only by Annelida but also by Arthropoda. Since Arthropods are Ecdysozoa and Annelids are Lophotrochozoa obviously segmentation evolved separately in these two groups. http://tolweb.org/Bilateria/2459.
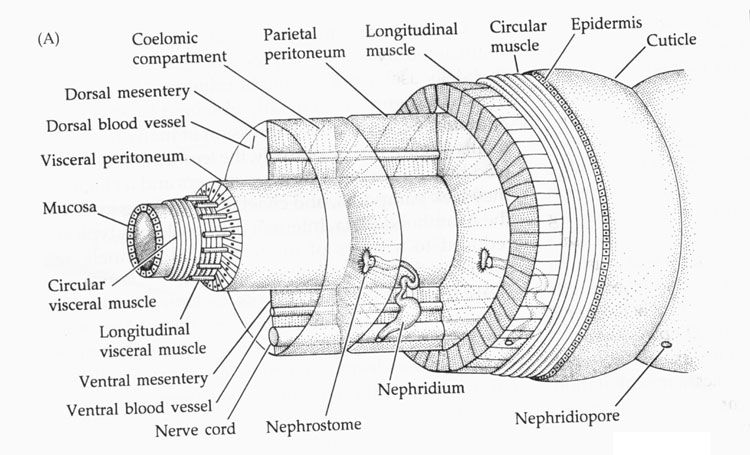
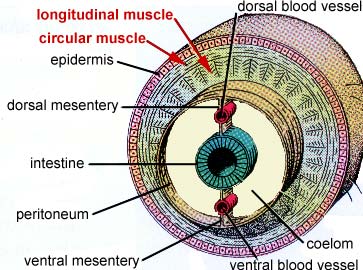
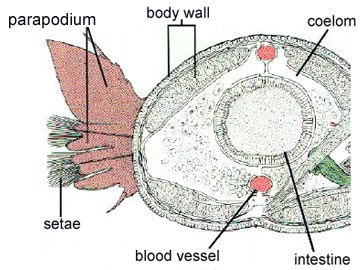
The annelid body plan is essentially a tube within a tube. This segmentation is both internal and external, and each internal segment of the body is separated by a tissue called septa, which partially divides the coelom. Segmentation is also called metamerism. Segments each contain elements of such body systems as circulatory, nervous, and excretory tracts.
Segmentation increases efficiency during development as it involves creating an entire body out of identical modules. It also aids locomotion because each segment acts independently of the other segments, giving the body greater flexibility.
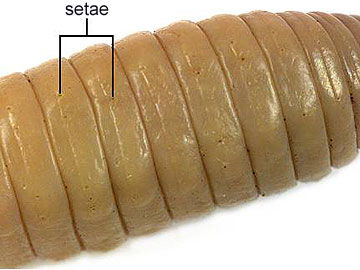
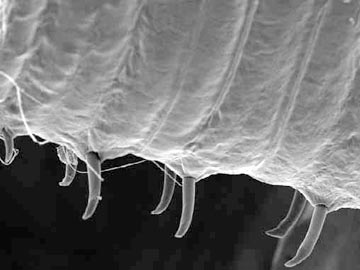
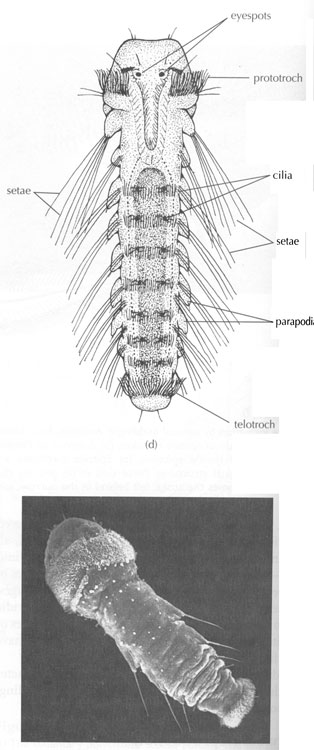
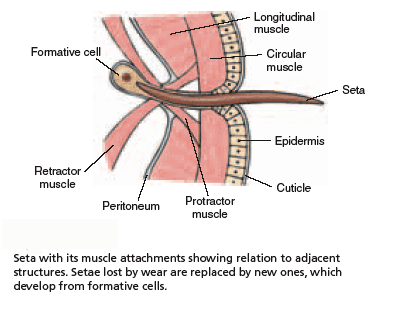
Besides being segmented, the body wall of annelids is characterized by being made up of both circular and longitudinal muscle fibers surrounded by a moist, acellular cuticle that is secreted by an epidermal epithelium. All annelids except leeches also have chitinous hair-like structures, called setae, projecting from their cuticle. Sometimes the setae are located on paddle-like appendages called parapodia
Muscles and locomotion:
Annelids have two sets of muscles that are used to contract and elongate the body.
Annelid worms are excellent examples of a hydrostatic skeleton in the form of a coelom in action.
-------------------------------------------------------------------------------------------
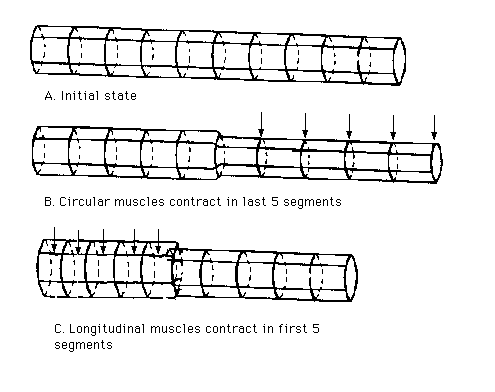
In earthworms, the hydrostatic skeleton or coelom is surrounded by sheaths of longitudinal and circular muscles. When the longitudinals shorten a short fat animal is produced, when the circulars contract a long thin one.
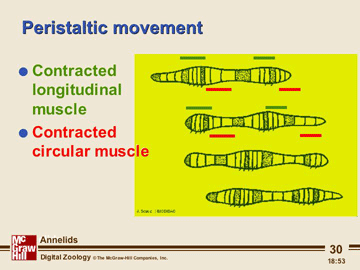
Locomotion begins with a contraction of the circular muscles in a limited region of the anterior end of the body. This contraction passes backwards down the body as a peristaltic wave, each wave being followed by a wave of contraction of the longitudinal muscle; . This continues in regular alternation.
At those regions where the longitudinal muscle is contracting the body surface bulges outwards and the setae are protruded. At these points, the worm can exert a thrust against the substratum. Where the circular muscle is contracting the body becomes thinner, and the setae are withdrawn. Here the segments extend forwards, aided by the thrust from the swollen regions. Thus the extension of the thinner parts of the body can be translated into forward movement.
Peristaltic movements of this sort can be used to drive the animal along and if, as if earth worms, some form of anti-slip device is incorporated into the design, considerable forward pressure can be generated. The importance of the thrust against something like a burrow wall and anti-slip device can be shown by placing the worm on a glass plate or other slick surface. Although the hydroskeleton again gives effective movement in a viscous media (a burrow), it proves ineffective for true locomotion on land.
_____________________________________________________
Earthworm movement is very well depicted in the following animation.
http://www.biology.ualberta.ca/courses.hp/zool250/animations/Earthworm.swf
These worms also use setae to anchor themselves within the burrow; contraction enlarges the body causing the setae to sink into the burrow's sides. The advantage of this can be seen everytime we witness a bird attempting to pull an earthworm from its burrow!
Leeches move across a surface by utilizing body muscles plus the anterior and posterior suckers. This mode of locomotion is much like that of an "inch-worm". Leeches can also swim by undulating the body.
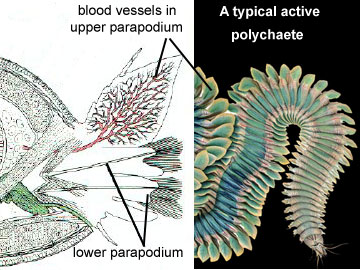
Polychaetes have several means of locomotion. When crawling slowly, they use the parapodia much like legs to propel themselves over their substrate. Thus, the parapodia have a dual function, the upper part being a respiratory organ, and the lower part a propulsive one. Muscles on either side of the body can contract out of phase, producing a rapid wiggling motion.
http://www.biology.ualberta.ca/courses.hp/zool250/animations/Polychaete.swf
Polychaetes have setae but do not utilize setae for locomotion. Sedentary worms may use hooked setae to anchor in their tubes. In some worms, the setae are defensive, penetrating the body of the attacker, breaking off, and causing irritation.
Organ systems
We will use the earthworm as our model animal, an Oligochaete (old phylum), belonging to the class or clade Clitellata, or Annelids with a clitellum, a swelling towards the head of the animal. The other major clade is the Polychaeta. With the exception of a few terrestrial species such as earthworms, the Oligochaetes mainly inhabit fresh water. Some species live freely in their aquatic environment, whereas others encase themselves in tubes. The earthworms live on land, but burrow deeply into damp soil. They may emerge at night or on rainy days, but are usually found within their burrows.

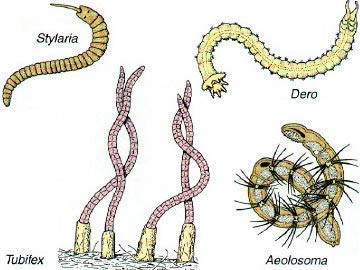
The many species of Oligochaete worms have a similar appearance. Except for difference in size, most of them resemble the common earthworm in external anatomy. Size however can affect the complexity and types of some of the systems observed.
Some diagrams of microscopic oliogochaetes.
Note in this photograph of a microscopic oliogochae, even segmentation is not as obvious as in larger forms.
All Oligochaetes bear bristles called "setae" on most segments. In the earthworms, setae are very short and can be best seen under the microscope.
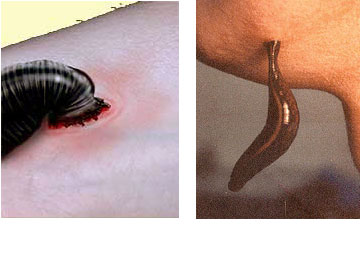
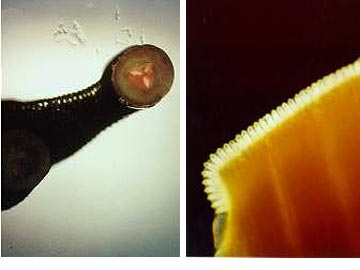
Leeches, ( Hirudinea), are also Clitellates. The Hirudinea resemble the oligochaetes in their major habitats (fresh water and land) as well as in lack of parapodia and reduction of specialized head structures that we will find in the Polychaeta. Leeches differ from the oligochaetes in their shape, by the absence of setae, and by the presence of suckers at both ends of the body. Clitellae may be present only during reproduction.
In the past, before the nature of disease was understood, leeches were used extensively in dubious medical treatment. Leeches are used in medicine today, but in limited, specific ways, including surgical procedures.
http://www.usatoday.com/news/health/2004-07-07-leeches-maggots_x.htm
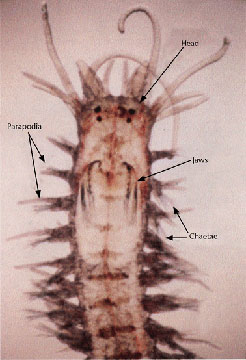
The polychaetes are marine worms, usually dwelling on sandy or muddy sea bottoms or along rocky coasts. Polychaetes differ from the Oligochaetes by the presence of parapodia and by a more specialized head region.
Many polychaetes live active lives: they swim, crawl or burrow near or on the ocean floor. Other polychaetes live in permanent burrows or tubes which they seldom, or never, leave. The diversity among polychaetes, especially segmentation specialization and head region is tremendous. About 80 percent of all known annelid species are polychaetes
Their delicate beauty have caused some species to be named after famous Greeks, nymphs and goddesses of Greek myth, such as Nereis (the common "clam worm") and Aphrodite (the "sea mouse")
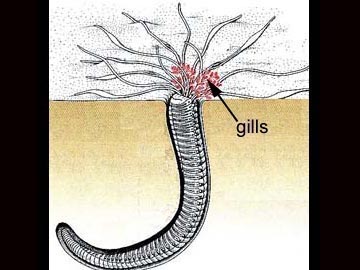
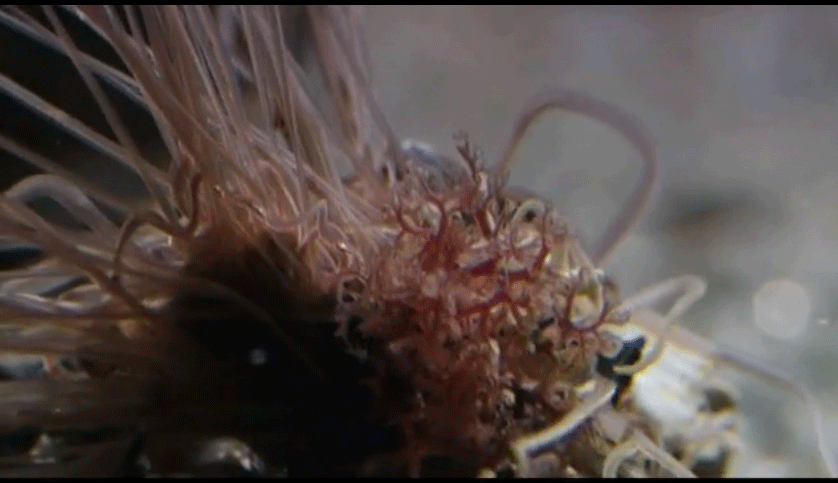
Respiration: Tiny blood vessels are abundant in the skin, which functions as the respiratory organ. Gas exchange occurs between the capillary beds on the body surface of the annelid and the environment. The oligochaetes and the leeches by and large exchange respiratory gases directly through the skin and have no special respiratory organs. For oligochaetes living on land, it is crucial that the body surface be kept moist to allow gas exchange to occur. Some aquatic annelids have thin-walled, feathery gills through which gases are exchanged between the blood and the environment. By contrast, the polychaetes have several different kinds of respiratory organs. The sedentary polychaetes may use their tentacles as respiratory surfaces or may have specialized gill structures protruding from their tubes.
The active polychaetes utilize parapodia for gas exchange. Each parapodium is divided into an upper and a lower section; it is the upper section that serves a respiratory function.
Digestive system:
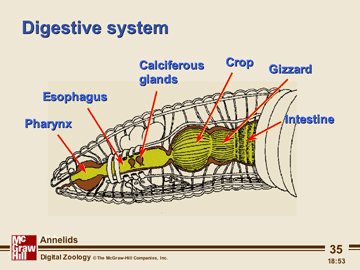
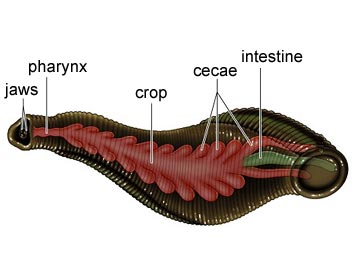
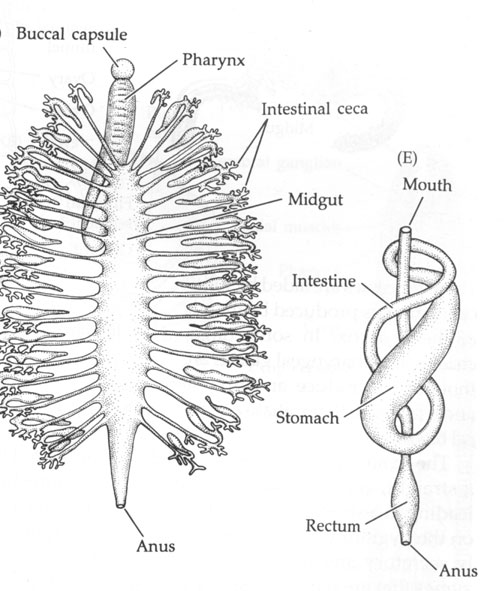
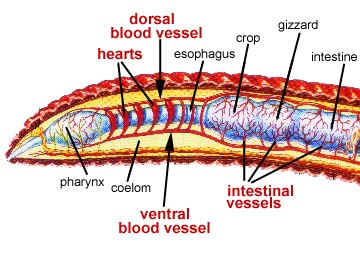
Annelids acquire food in ways specific to their diets which vary widely. However, most annelids have a muscular pharynx leading into a straight tube which serves as esophagus, stomach and intestine. Variations in this structure are associated, again, with diet.
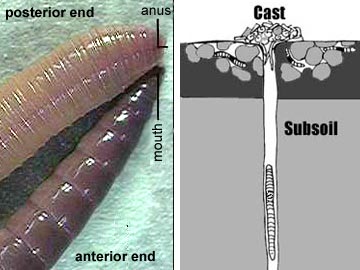
Terrestrial oligochaetes (the earthworms) consume soil. After entering the mouth, the soil is sucked into the pharynx and passes through the digestive tract where small particles of dead organic matter within the soil are digested. Parts of the esophagus are modified into a crop for storing food and a gizzard lined with hardened cuticle for grinding fool. Calciferous glands are associated with the esophagus and believed to function in regulating blood pH by controlling the concentration of calcium ion. The intestine is very long, as is necessary to absorb sufficient nutrients from the soil. The surface area for absorption is further increased by a dorsal infolding of the intestinal wall. Undigested matter is eliminated as castings through the anus at the posterior tip of the worm.
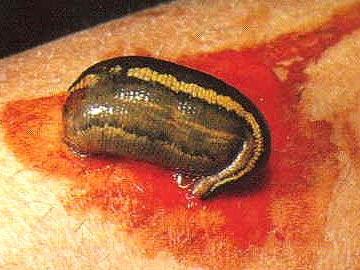
Many leeches are parasites.
http://shapeoflife.org/video/annelids-leeches
They attach to a host, usually a vertebrate animal, and feed on its blood. Fish, amphibians, mammals, and even birds can serve as hosts. Parasitic leeches locate a suitable host by sensing body heat. They attach to the host by the anterior sucker which surrounds the mouth. Some leeches have sharp jaws with which they make a three-way incision into the flesh of their victim, then consume the blood as it flows from the wound. Other leeches lack sharp jaws and instead of cutting a wound, they secrete a powerful enzyme that digests a hole in the skin of their host thereby giving them access to its blood. The digestive tract of leeches is more complex than those of oligochaetes and occupies a larger portion of the coelomic cavity. This is facilitated by loss of the internal partitions between segments in leeches. Digestion and absorption of food and nutrients occurs within the intestine of both predacious and sanguivorous species with only the predacious species additionally utilizing the crop for these functions. In sanguivorous species symbiotic bacteria aid in the digestion of blood meals by producing enzymes that aid in the breakdown of blood. Several species of bacteria have been found to colonise leeches living on blood such as Hirudo medicinalis. Digestion in sanguivorous leech species may take weeks or even months due to the reliance on endosymbiotic bacteria. Predatory species utilizing their own enzymes digest their food within a few days
Polychaetes vary in their food habits.
Some polychaetes are active predators and accordingly have jaws. The jaws are attached to the pharynx and can be retracted into the pharynx cavity or everted through the mouth to capture prey.
Some tube dwellers also feed on other animals or plants found near by.
http://shapeoflife.org/video/annelids-diopatra-tube-dwelling-worm
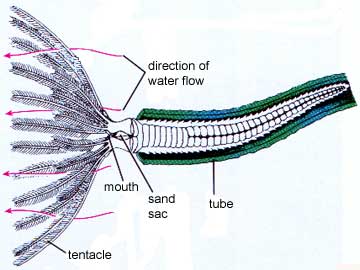
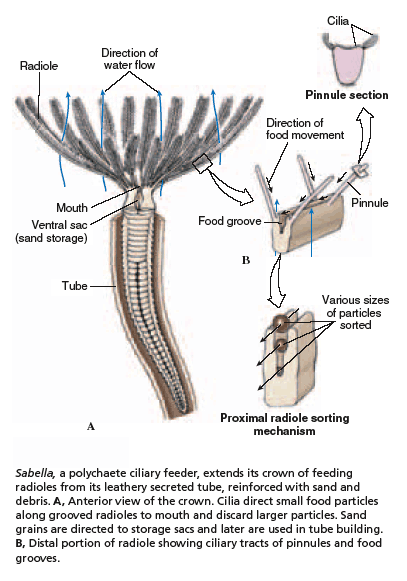
Sedentary polychaetes are often filter-feeders. Tube-dwellers, such as the fan worms, utilize their large feather-like tentacles to filter small food particles from the water. Tubes vary in complexity of construction. Some are made up of sand or mud particles cemented together with digestive secretions. In others the worms also contribute calcium carbonate or proteins and polysaccharides.
Polychaete that live in burrows create a current of water flow through their burrow and feed on food matter brought in with the water.
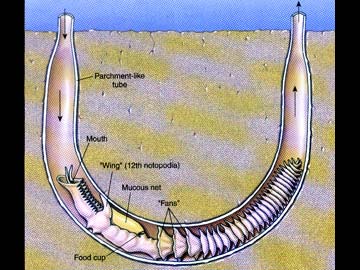
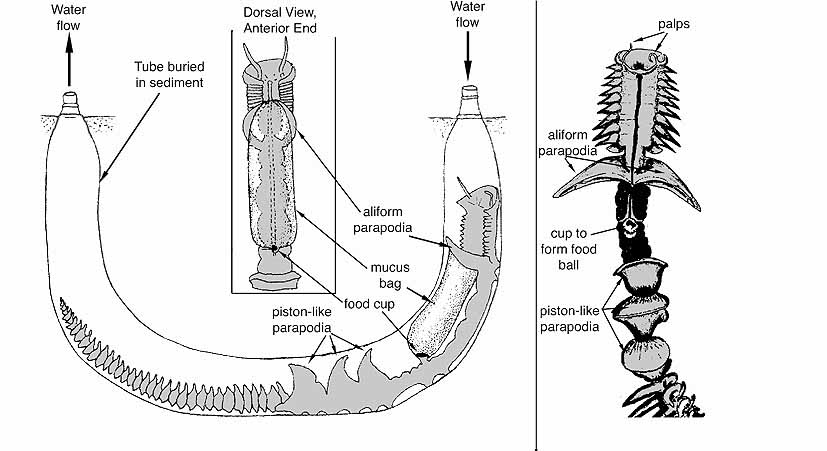
Example: Chaetopterus
The piston-like parapodia pump water through the tube from anterior to posterior. A mucus bag held between the aliform parapodia and connected to the food cup acts to filter the water passing through the tube. The food cup continually rolls up the mucus bag and and when the bag reaches a certain size it is is detached and passed forward to the mouth
Circulation
Annelids have a closed circulatory system. This means that the blood is contained entirely within hearts and blood vessels and does not drain into the body cavities. In annelids, there are two major blood vessels which run down the length of the body and are connected to one another in most segments by interspersed capillary beds that lie along the digestive tract. In the anterior end, the two longitudinal blood vessels are connected by larger vessels that contract rhythmically, thus serving as "hearts".
The close association of the circulatory system with the digestive tract allows for absorption of nutrients from the gut into the blood, which then transports them to the various parts of the body. In oligochaetes, oxygen diffusing through the body wall can be picked up from the coelomic fluid by the blood and transported to wherever it is needed, with carbon dioxide following the reverse route. Thus the coelom also serves as a transport system, albeit to a limited extent.
Excretion
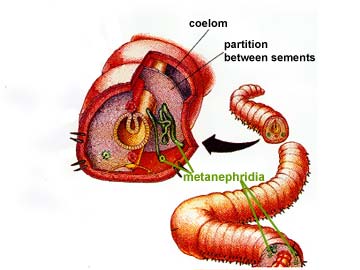
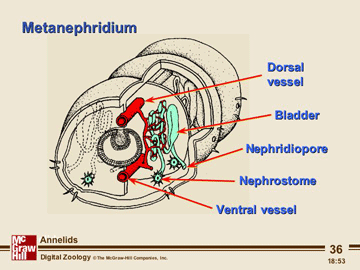
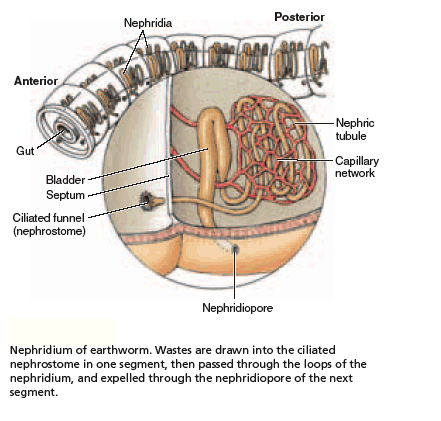
The nitrogenous waste products of metabolism diffuse from the body's cells into the fluid of the coelom, which thus plays a role in waste transport. The coelomic fluid is slowly drained into small tubular organs, called metanephridia (singular metanephridium). As the fluid passes through the metanephridia it can exchange some materials with the network of blood vessels that are intertwined around the tubule. Accordingly, the content of the fluid may become somewhat modified.
http://www.biology.ualberta.ca/courses.hp/zool250/animations/Excretion.swf
The ability of annelids to modify their excretory products allows for osmoregulation to occur. For example, certain polychaetes can live in estuaries where the salinity of the water fluctuates widely. Finally, the fluid with its contained nitrogenous wastes exits the body through tiny pores in the body wall.
Nervous system
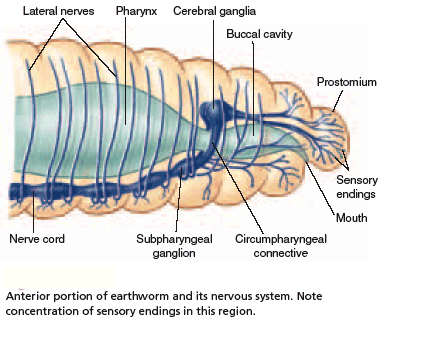
The basic annelid nervous system consists of a single or double nerve cord running along the ventral side of the body with an enlarged region (ganglion) in each segment. Active annelids, such as the earthworm, have larger ganglia in the head region that serve as simple "brains".
The segmentally organized nervous system of an annelid, with the longitudinal connectives and ganglia fused along the midline. There is an elongated ganglion for each segment.
Annelid sense organs then are simple and vary according to life style of the worm. Oligochaetes do not have discrete sensory organs, although the skin, particularly in the anterior region, has cells with sensory functions. Some are light-sensitive, others chemosensory and still others are sensitive to touch.
The active polychaetes have a well developed head region bearing tentacles and palps that are sensitive to chemicals and touch. The head also bears eyes that range from simple eyespots to large conspicuous eyes on stalks.
Reproduction:
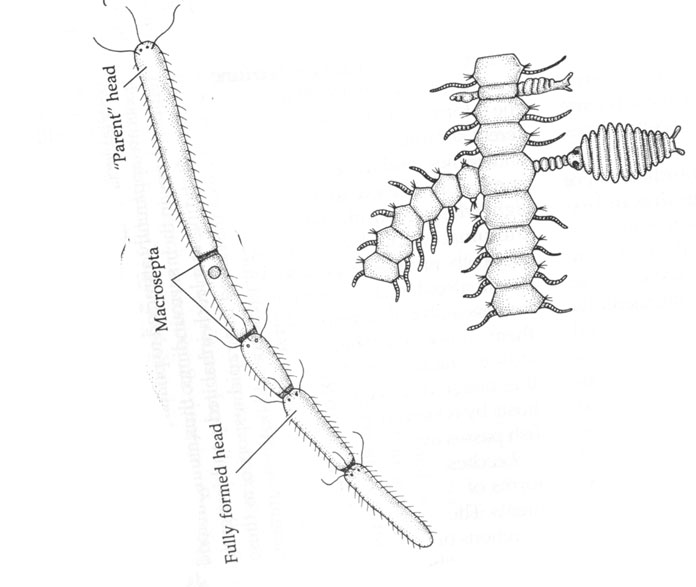
Asexual reproduction by fission is a method used by some annelids and allows them to reproduce quickly. The posterior part of the body breaks off and forms a new identical worm.
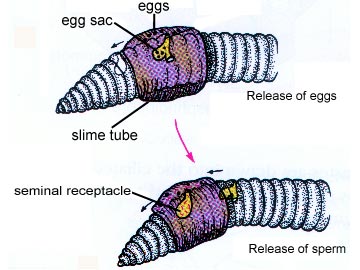
Oligochaetes are hermaphroditic and practice a form of mutual copulation. The sperm from each worm's body are transferred to the other worm (some species have penises) and then move to the spermatheca in the other worm, where it is stored until fertilization.
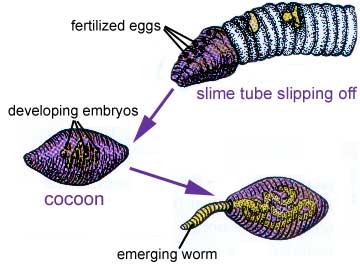
A complex cocoon is eventually secreted by the clitellum into which eggs and sperm are secreted, so fertilization is external. The worms separate and form cocoons; the cocoon moves forward, picking up eggs at the 14th segment; at the 9th and 10th segments it picks up the sperm deposited by the other earthworm. The embryo develop in this "cocoon" which will be shed by the worm. There is no larval form in oligochaetes, even in marine species.
Leeches are hermaphroditic and most of them have reproductive habits similar to earthworms. They have a clitellum which produces a cocoon that slides off the animal, collecting the gametes as it does so.
However, other leeches have a more innovative way of reproducing. One leech deposits a packet of sperm onto the body of its partner. The sperm then bore through the skin and fertilize the eggs. In this case, fertilization in truly internal!
Although most leeches deposit their cocoons, then leave, a few species carry the cocoons with them until the young leeches emerge.
Polychaetes have separate sexes and fertilization is external. Polychaetes are unique in not having well-defined gonads. Rather the gametes are produced by germ cells located in the lining of the coelom or in the walls of the septa between segments. Like most aquatic invertebrates, polychaetes reproduce by shedding their gametes into the water, and fertilization occurs in the aqueous environment. As noted previously, some polychaetes can also reproduce asexually by budding off individual body segments that then grow into complete worms.
Development
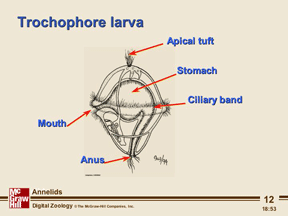
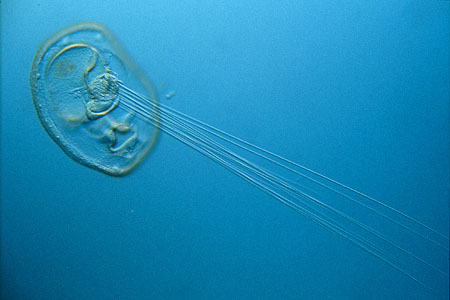
The early embryonic development of all annelids is essentially similar. Though eggs can range in size from less than 0.05 mm in diameter to more than 1 mm, they all show a sequence of spiral cleavage to the 64-cell stage. The tissue layers are formed from a few well-defined cells in the blastula. For example, all of the future mesoderm is formed from the descendants of the 4d cell. In some polychaetes the zygote develops into a free-swimming trochophore larva. Other polychaetes produce larval forms that are somewhat different. The trochophore larva is an important feature, because it links the annelids to the mollusks, of which some marine forms have a trochophore larva.
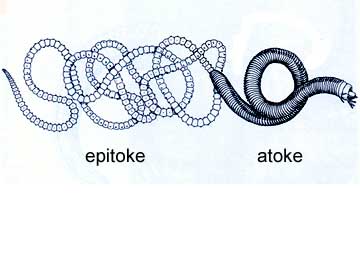
A few polychaetes have a unique way of maximizing the likelihood that their eggs will be fertilized. In Paolo worms, special gamete-filled segments (epitokes) break off the body and swim to the surface of the sea where they release large numbers of gametes. Some text consider epitokes to be asexual produced individuals, although in some worms transformation of old segments rather than the production of new ones is involved. Worms that reproduce in this way often spawn at the same time and place. Some species produce large swarms at the water surface where the worms release their gametes.
Male palolo worm
Female Palolo worm
Visit the websites below to learn how the Palolo worm or more correctly their epitokes reproduce.
https://www.youtube.com/watch?v=5zR3CjhpABU
https://www.youtube.com/watch?v=VlsHiu04SEI