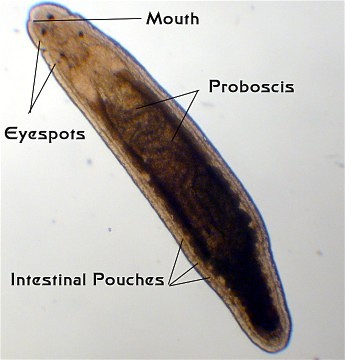
Superficially, the ribbon worms (Phylum Nemertea) look like "unflat" flatworms. The number of species is estimated to be between 900 and 1400. Cilia on the bottom surface move the animals in a gliding locomotion.
A common feature of many of the ribbon worms is the extremely thin and elongated, unsegmented body. Some can reach a length of 30 meters with a body diameter of only a few millimeters.

They are more complex than flatworms. They have a complete digestive tract, with mouth and anus. A complete digestive tract is considered an advantage, since the one-way flow of food allows for regional specialization.
They also have a closed circulatory system.
One of their unique characteristic is their probosis.
To elaborate:
The internal anatomy of these animals is complex.
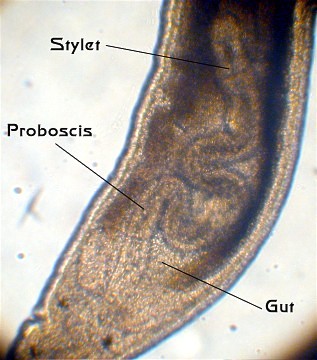
GI track: The gut is a complete gut with sequential digestion occurring from front to back in the gut. Unlike the gut found in flatworms, which has only one opening, the gut in ribbon worms has an anterior mouth and a posterior anus. The mouth may be found either at the front of the body, or on the bottom of the body a short distance behind the front end. The gut has quite a few internal divisions, but they don't correspond closely to what is seen in many other animals. Behind the mouth is a short esophagus, often with a large pouch or caecum extending from it. Located just after the esophagus is the relatively small stomach, and then a long intestine extends almost all the way to the end of the worm. The intestine has many side pouches, referred to, collectively, as intestinal caecae. It appears that much of the digestion occurs within the cells lining the various pouches rather than in the gut cavity. A short hindgut terminates in the anus. The gut is embedded in the body musculature, and no body cavity is found surrounding the gut.

Proboscis: The single structure that sets these worms apart from all other worms is their proboscis. Basically, the term "proboscis" simply means a large, evident and sometimes mobile nose.
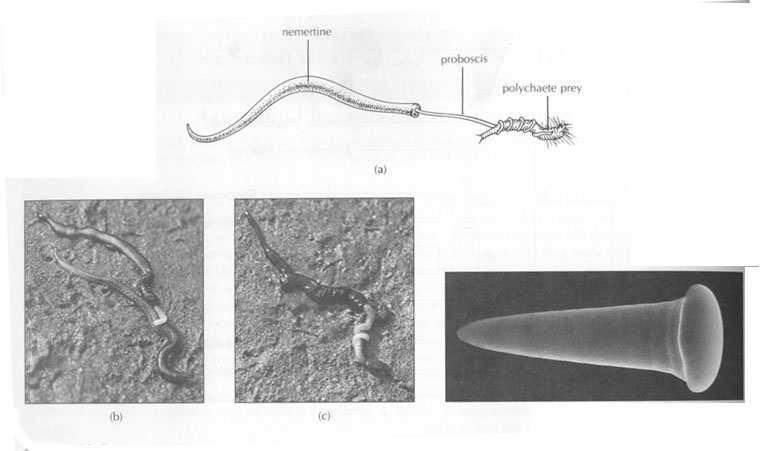
Here the proboscis is a rapid-strike prey capture organ. The nemertean proboscis is normally retracted into an internal sheath or pouch found above the worm's gut. The proboscis sheath is a closed impermeable tube and is filled with fluid. When the animal senses a prey organism nearby, a circular muscle layer around the proboscis sheath rapidly and vigorously contracts. This contraction forces the fluid from the proboscis sheath into the proboscis and, in the process, literally turns it inside out, blowing it out of the proboscis sheath. The proboscis will rapidly (within a second or so) wrap itself around the prey, which is then drawn to the mouth and eaten. Nemertean muscles, particularly the proboscis retractor muscle, show unusual extensibility, sometimes achieving a 50-fold change in length. Probosis diversity.

Circulation:
The ribbon worms possess a closed circulatory system. The blood always is found with vessels. There is no heart, nor are there valves within the system. Blood simply sloshes around within the vessels and is moved by muscle contractions throughout the body. The blood may or may not contain hemoglobin, but if it does, the hemoglobin is not found in cells. Uniquely among invertebrates blood vessels are formed by splitting of longitudinal mesodermal tissue. Vessels are situated deep beneath the body wall musculature and are probably not implicated in oxygen transport. There is some variations in the plan of the blood system. There is a blood vessel loop around the brain region and lateral blood vessels which extend down the length of the body on either side. Frequently vessel elaboration occurs around the foregut or alongside excretory tubules. The mid-dorsal vessel usually penetrates the rhynchocoel near the brain; the proboscis has no direct blood supply.
Excretory system consists of protonephridia. In many species these are closely associated with the circulatory system and so probably do function in the processing of nitrogenous wastes. There is some evidence that as in Platyhelminthes they also function as osmoregulatory organs.
They have a large bilobed brain with a major nerve running around the front of the gut, in the region of the throat. Instead of a single spinal cord, they have paired major nerves running the length of the body on either side. Additionally, large nerves run from the brain to the proboscis. They typically have simple eyespots, sometimes as many as several dozen. These eyespots cannot form images, but in the most sophisticated receptors can detect changes in light intensity and the angle of light impingement. Additionally, they have several well-enervated organs in the head region that are thought to be mostly chemosensory.
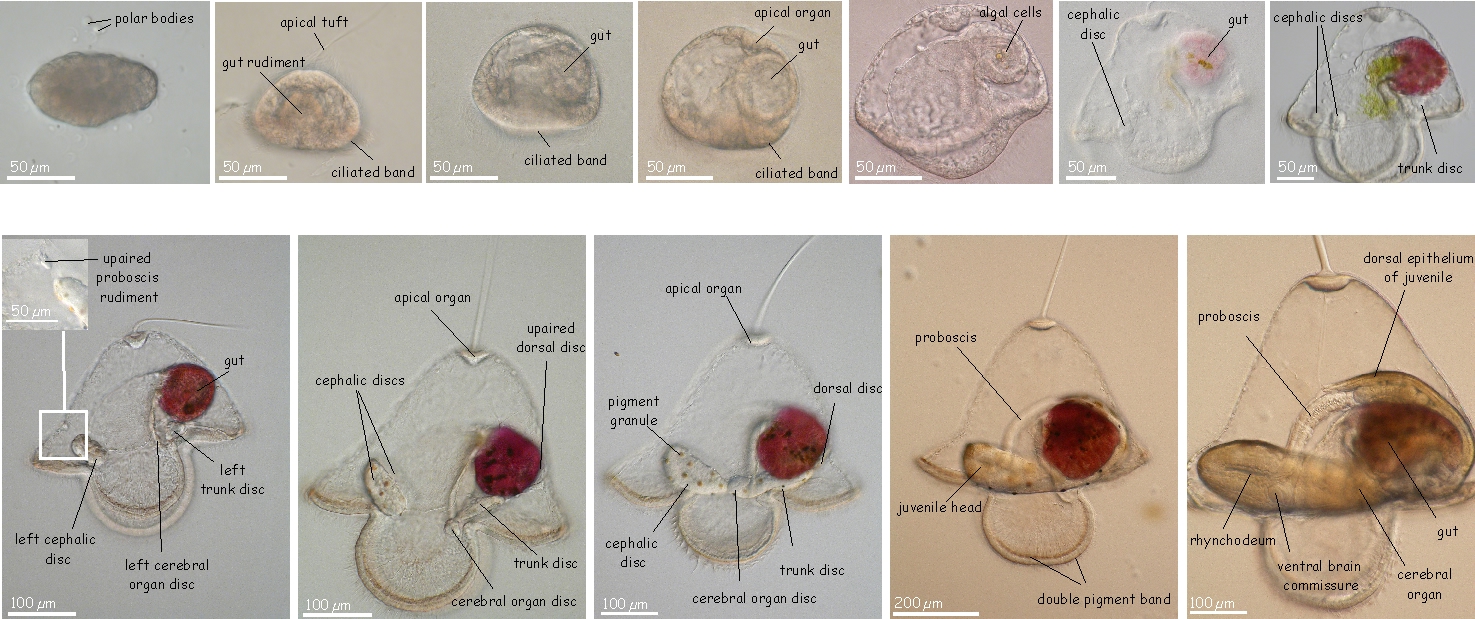
Their reproductive systems are simple to the point of being rudimentary. The sexes are separate, and the gonads are found as masses of egg or sperm producing tissue between the intestinal pouches. When the animals are gravid, and "the time is right," temporary openings form to the outside and spawning occurs. In a few nemerteans, such as Cerebratulus species, a specialized hat-shaped larvae, called a "pilidium" occurs. This larva develops and then undergoes a metamorphosis into a juvenile worm. The interest in this type of development is due to the presence of imaginal discs (think insect pupae). These discs are embryonic tissue but programmed to form specific adult tissues and organs at metamorphosis. In most nemerteans, the embryo develops directly into a small worm, and has no larval form. They also develop asexually by fragmentation.
Development of larvae below from faculty.washington.edu/maslak/research.shtml
This phylum includes, mainly marine, species. Most nemerteans are predators, but some are scavengers and others are herbivorous. They are often found in and among seaweeds, rocks, mussel and barnacle beds, or buried in mud, sand, or gravel substrates. Some nemerteans actively track prey by following their chemical trails, - common prey items are polychaetes , snails , small crustaceans , and mollusks. Usually they feed on scavenges and dead animals. Food is located by chemical signals or simply by pure chance. Even species equipped with eyes do not necessarily use them to find food.
Several species of some specialized nemerteans are commonly found inside the mantle cavities of large clams. Although it is widely thought that the clam-dwelling nemerteans are parasites, in fact, they eat small crustaceans and worms found in the mantle cavities. They are probably best considered to be beneficial symbionts that clean the mantle cavities of pests.
Some of the truly parasitic forms live on crabs and are egg parasites. In North America, there are species that live on the outside of the crab's carapace. They eat the eggs of the crabs; each worm can eat an egg or so a day. An infestation of several dozen worms can effectively eat the entire yearly spawn of the crab.
No one really knows how this group, often being so conspicuous defends itself against predators, but some work point to bacterial symbionts that produce defensive chemicals. Your books documents the production by some species of a powerful neurotoxin.
No known fossil can with certainty be related to nemerteans. Evidence from early development (e.g. spiral cleavage) and from small subunit ribosomal DNA relates them to coelomate protostomes. They should probably be placed among the Lophotrochozoa. Both the rhynchocoel (effectively a restricted, localized coelom) and the blood vessels have their origin in mesodermal splitting, somewhat resembling the formation of the annelid coelom. Some people suggest that nemerteans are regressed coelomates, but regression followed by radiation into a phylum with more than 1000 species is considered highly improbable. Coelom-like cavities have arisen several times separately during evolution. Nemerteans may have developed from an early acoelomate stock and remained a distinct clade with its own evolutionary history through time.